Род больших кошек, Panthera Oken, 1816
1816. Panthera*. Oken. Lehrb. Naturgesch. 3, 2, p. 1052. Felis pardus. Linnaeus.
* (Название не вполне удовлетворяет правилам номенклатуры, однако используется широко. Заслуживает сохранения как nomen conservandum.)
1816. Tigris. Oken. Ibid. 3, 2, p. 1066. Felis tigris Linnaeus.
1816. Leo. Oken. Ibid. 3,2, p. 1070. Febis leo Linnaeus.
1829. Leo. Brehm. Isis (Oken). p. 637. Felis leo Linnaeus.
1843. Tigris. Gray. List Mamm. Brit. Mus., p. 40. Felis tigris Linnaeus.
1868. Pardus. Fitzinger. Sitz.-Ber. K. Ak.Wiss. Wien. 58, 1, p. 459. Felis pardus Linnaeus. (В. Г.).
Размеры крупные и очень крупные - самые крупные в семействе.
Общий облик вполне типичный для всех кошек. Ноги у большинства невысокие, толстые, туловище вытянутое, иногда сильно. Крестцовая область не высокая, и линия спины кзади не поднимается - она прямая или высота в плечах (в холке) больше, чем в крестце, и линия спины слегка поката кзади (рис. 30). Хвост длинный, не менее половины длины тела, у некоторых немного больше. Голова умеренной величины или крупная, несколько вытянутая, с относительно удлиненной лицевой частью и более или менее прямой или слабо дугообразной верхней линией профиля. Уши относительно маленькие, короткие, с тупой закругленной вершиной, широко расставленные. У самцов льва и у зверей в зимнем наряде они слабо выдаются из меха. Удлиненных волос (кисточки) на вершине уха не бывает. Глаза относительно маленькие, зрачок круглый.
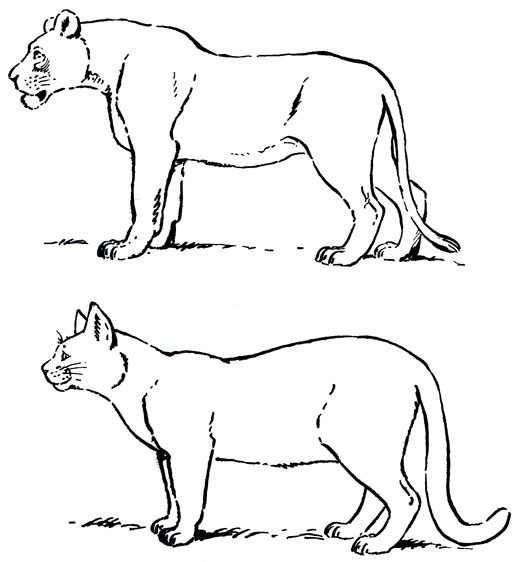
Рис. 30. Абрис профиля тела большой, p. Panthera и мелкой, p. Felis, кошек. Характерная приподнятость крестцовой области у мелких кошек. Львица и домашнаяя кошка (рис. Н. Н. Кондакова)
Волосяной покров у большинства видов по всему телу ровный, и хвост равномерно опущен по всей длине. Иногда волосы на щеках удлинены (баки), у одного вида (лев) в передней части тела самцов хорошо развита грива, и хвост имеет на конце кисть из удлиненных волос. Ноги сильные, с широкой мощной лапой, особенно передней, когти большие, очень острые и круто изогнутые, полностью втяжные. На концах всех пальцев с внутренней и с внешней стороны когтя есть кожная складка, полностью скрывающая втянутый коготь. Межпальцевые перепонки большие, доходят до конца пальцевых подушечек (рис. 278). У большинства волосяной покров короткий и не густой, лишь у крайних северных форм некоторых видов зимой он бывает густ и относительно длинен. Окраска одноцветная или с черными поперечными пололасти у мелких кошек. Львица и домашняя кошка (рис. Н. Н. Кондакова) сами или по светлому фону разбросаны черные пятна как одиночные, так и Образующие кольцевые фигуры. У большинства видов на тыльной стороне уха большое белое пятно. Сосков 2 или 3 пары.
Череп крупный и тяжелый с массивными костями, с толстыми и мощными скуловыми дугами и сильно развитыми буграми и гребнями. В частности, длинен и очень высок сагиттальный гребень, сильно выдающийся назад, и Сильно развит затылочный. Верхняя линия профиля довольно прямая или лишь слабо и равномерно выпуклая, межглазничная область приподнята относительно слабо, и линия профиля кзади от нее понижается полого. В целом череп выглядит уплощенным. Базикраниальная ось его (ось основания мозгового черепа) почти горизонтальная и не образует выраженного угла с базифациальной. В месте перехода носовой области в межглазничную на верхней части черепа впадин (вдавления) нет.
Черепная коробка относительно небольшая, у некоторых очень небольшая, вытянутая, как бы сильно сдавленная жевательной мускулатурой, крепящейся к сагиттальному гребню. Заглазничное сжатие сильное, иногда его ширина равна или даже меньше ширины межглазничного. Лицевая часть черепа очень мощная - сильно развита как в ширину (в скуловых дугах и области клыков), так и в длину. Считая от поперечной линии, проходящей по надглазничным отросткам, лицевая часть больше или не меньше мозговой в целом, или без нависающей над затылком части сагиттального гребня.
Между засочленовным отростком и слуховым барабаном довольно большое расстояние - приблизительно равное ширине барабана. Слуховые барабаны относительно малы, не вздуты, с вентральной стороны уплощены и толстостенны (у барса невелики, но выпуклы); их внутренняя камера (энтотимпанальная часть) значительно больше наружной (эктотимпанальной, или собственно тимпанальной), и перегородка между ними лежит близко к наружному слуховому проходу. В подъязычном аппарате (рис. 31) третий сверху элемент (epihyale) не окостеневший и представляет собой эластичную связку, которая допускает значительную подвижность гортани (см. далее). На базиокципитальной части затылочной кости вплотную к костному барабану кпереди от яремного отверстия глубоких вдавлений нет или они едва обозначены.
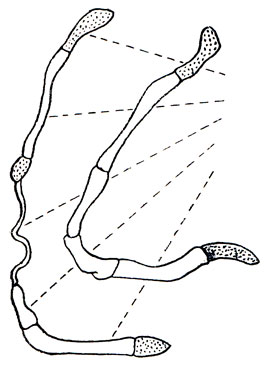
Рис. 31. Дужки подъязычного аппарата (схема) рода больших кошек, Panthera Oken (слева), рода мелких кошек, Felis L. и гепардов, Acinonyx (рис. Н. Н. Кондакова, по X. Хеммеру)
Зубная система полная - второй предкоренной (первый в ряду) верхней челюсти обычно имеется. Зубы очень сильные, клыки относительно коротки, но мощные, с широкой основной частью.
Половой диморфизм у большинства видов выражается только в больших размерах самца, частью более сильном развитии гребней и т. п. на черепе, возрастной деморфизм в окраске не выражен. У одного вида (лев) самец резко отличается развитием гривы на передней части туловища и удлиненных волос местами на груди и брюхе; при одноцветной окраске взрослых новорожденные и молодые имеют окраску пятнистую. Сезонного деморфизма в окраске нет; у северных или горных подвидов некоторых видов зимний мех гуще и длиннее.
Хищники, добывающие главным образом крупных млекопитающих (копытные). Добыча обычно превышает собственные размеры зверя, иногда в несколько раз. Охота скрадом и подстереганием на тропах, у водопоев и т. п. Ночные, за одним исключением одиночные звери, причем самец не принимает участия даже в выкармливании потомства. Один вид (лев) всегда держится и охотится небольшими стаями, по-видимому, представляющими собою семейные группы по типу волчьей стаи (см. т. II, ч. 1). Обитатели равнинных и горных лесов, иногда открытых безлесных гор, тростниковых зарослей, один вид - открытых степей (саванн) и полупустынь.
В связи с особым строением подъязычного аппарата (см. выше) гортань с ее голосовыми связками обладает большой подвижностью, призывный звук представляет собой громкий грубый рев ("ревущие кошки" - "мяукать" большие кошки не способны). Удовольствие и радость "мурлыканьем" не выражают, или "мурлыкают", по-видимому, только при выдохе. Едят, главным образом, лежа на брюхе и опираясь предплечьем и локтями на субстрат, куски добычи отрывают рывком головы вверх (рис. 32).

Рис. 32. Обычная поза при еде большой кошки, р. Panthera - предплечье горизонтально на субстрате, и мелкой кошки, p. Felis - локоть высоко поднят (рис. Н. Н. Кондакова)
Ареал рода (восстановленный) (рис. 33) занимает Африку, крайний юго- восток Европы, Азию, кроме северной части, и Южную и Центральную и самый юг Северной Америки. В Южной Америке ареал занимает всю северную и частью среднюю часть материка, на юге включая Перу, Боливию, Парагвай и юг Бразилии приблизительно до Сан-Пауло, а в Аргентине-провинции Сан-Луис и даже Ла-Пампа (немного севернее 40° с. ш.). На севере ареал охватывает всю Центральную Америку приблизительно до меридиана Вера-Крус в южной Мексике и далее, обходя с востока и запада Мексиканское нагорье, вдоль берегов Тихого океана и Карибского моря двумя длинными мысами протягивается с одной стороны в южный Техас, с другой-в штаты Калифорния, Аризона и Новая Мексика. Собственно п-ов Калифорния и все острова западного полушария в ареал не входят.
Африку ареал занимает всю, в Европе-южные части Балканского полуострова. В Азии южная граница охватывает Аравийский полуостров, идет по южному побережью материка, включая Цейлон, Малакку, Суматру, Яву, Бали и о-ва Кангеан (к северу от Бали). На востоке граница идет также по побережью материка, причем на Хайнане*, Тайване, Японских и Шантарских островах видов рода нет. На Сахалине звери бывали заходом (тигр). На севере в Азии ареал захватывает Приамурье, Забайкалье, Алтай и Кашгарию, Восточный и Южный Казахстан и Среднюю Азию, Иран, Кавказ и Малую Азию (подробности см. ареал тигра и барса). За историческое время ареал местами в связи с истреблением зверей значительно сократился (Европа, Средняя Азия, Сибирь, части Африки и Южной Азии и др.) и продолжает сокращаться местами очень быстро.
* (У В. Мазака (1965) на карте ареала тигра (стр. 33) указан (стрелка) заход на Хайнань. Это явное недоразумение: автор, очевидно, имел в виду старые сведения о случае захода тигра с материка на о-в Амой (5 миль от материка), приведенные, в частности, у Аллена (1938). О Хайнане в литературе (также и у Мазака, 1965) никаких положительных сведений нет.)
Виды рода принадлежат к наилучше изученным видам семейства, прежде всего в отношении морфологии, но также и этологии*. Разногласия между систематиками касаются лишь вопроса об относительной прогрессивности отдельных видов и выделения подродов (см. ниже).
* (Литература, посвященная этой группе кошек, очень обширна. К основополагающим относятся работы Северцова, 1858; Покока, 1917 и др., Хальтенорта, 1936, 1937 и др.; Вейгель, 1961 и др. и сводка Хеммера, 1966 (лит.).)
Как уже сказано выше, есть основания считать род крупных кошек в кругу родов семейства менее специализированным. Во всяком случае, сравнительно с другими группами семейства (разные группы видов в p. Felis), в которых "кошачий" тип специализации достигает своего наиболее полного развития и выражения, представители этого рода по структуре черепа представляют менее специализированный, более нейтральный "хищнический" тип. Возможно, что некоторые черты видов p. Panthera вторичны (алломатрия).
Род больших кошек появляется в раннем плиоцене Евразии. В раннем плейстоцене Евразии он был представлен особыми видами (P. arvernensis, P. cristata), иногда принимаемыми за предковые формы льва и тигра. Широко распространенный на протяжении плейстоцена пещерный лев, или "тигролев" (P. spelea) - самостоятельный вид, с которым современные львы и тигры, очевидно, непосредственной генетической связи не имеют. В Новом Свете род известен лишь в плейстоцене (гигантский ягуар Северной Америки, P. atrox).
В роде 4 современных вида - тигр, P. tigris Linnaeus, 1785, лев, P. leo Linnaeus, 1758, барс ("леопард"), P. pardus Linnaeus, 1758 и ягуар, Р. onca Linnaeus 1758, т. е. 11,5% видов семейства.
Все они составляют морфологически и филогенетически весьма тесную группу, но взгляды на соотношения видов довольно различны. Некоторые (Покок, 1929, 1930, 1941) подродов в роде не выделяют или, принимая в роде большее число видов (Хальтенорт, 1936-1937) относят перечисленные виды к одному подроду, другие (Хеммер, 1966) принимают два подрода, противопоставляя тигра, как наиболее примитивную форму, остальным или каждый вид выделяется в особый подрод (Эллермэн и Моррисон-Скотт, 1956, 1966; см. синонимику). Большинство систематиков рассматривает льва как наиболее примитивную форму рода и видит в тигре ближайший к нему вид. Предложено, однако (Хеммер, 1960), по сумме морфологических и этологических особенностей считать наиболее примитивным тигра, а льва рассматривать как наиболее специализированную форму рода. Здесь принимается чисто морфологический ряд лев - тигр - барс без выделения подродов. Наибольшее число видов (3) свойственно Старому Свету - 1 Африке и Азии (барс - "леопард"), 1. Африке, Азии и Европе (лев), 1 чисто азиатский (тигр). В Новом Свете лишь 1 вид (ягуар).
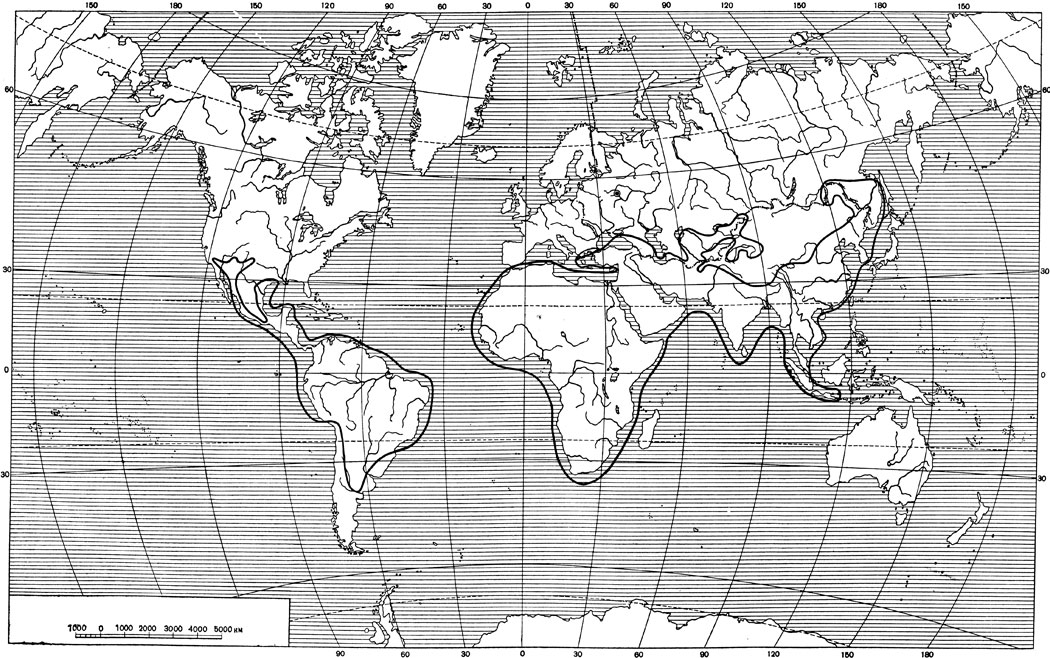
Рис. 33. Восстановленный ареал рода больших кошек, Panthera. В. Г. Гептнер
Одни из самых ценных спортивных охотничьих животных, регуляторы численности копытных, местами истребители вредителей сельского хозяйства (тигр - кабан, африканский барс - павианы), имеют значение и как пушнина. Местами вредят животноводству, представляют опасность и для человека.
В фауне СССР 3 вида - лев, P. leo Linn, 1758, тигр. P. tigris Linn, 1758, и барс ("леопард"), Р. pardus Linn., 1758. Это составляет 75% от числа видов рода, около 11,5% видов семейства и около 1% от числа видов фауны СССР. Лев истреблен полностью еще в X веке.
Ареал занимает южные горные и лесные, местами полупустынные и пустынные части страны, и в последние десятилетия сильно сократился. Лесные и горные формы, обитатели тростниковых зарослей. Ввиду малочисленности практическое значение очень невелико и редко бывает отрицательным. Один вид (тигр) находится под полной охраной; как и барс, представляет большую ценность для зоологических садов. (В. Г.).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://mur-r.ru/ 'Библиотека о кошачьих'