Генетическая терминология
Как установил Г. Мендель, каждый признак, присущий родителям и потомству, обусловлен факторами, передаваемыми потомству генами.
Установлено, что ген с биохимической точки зрения представляет собой часть молекулы дезоксирибонуклеиновой кислоты и кодирует один из передающихся по наследству признаков.
Различные формы одного и того же гена называют аллелями. Например, у кошки ген, характеризующий длину волосяного покрова, включает аллель (признак), определяющий короткую шерсть, и аллель, характерную для породы длинношерстной. В соответствии с фенотипом каждого индивида аллели (признаки) могут быть доминантными (их обозначают прописными буквами - А, Б и т. д.) и рецессивными (их обозначают строчными буквами - а, б и т. д.).
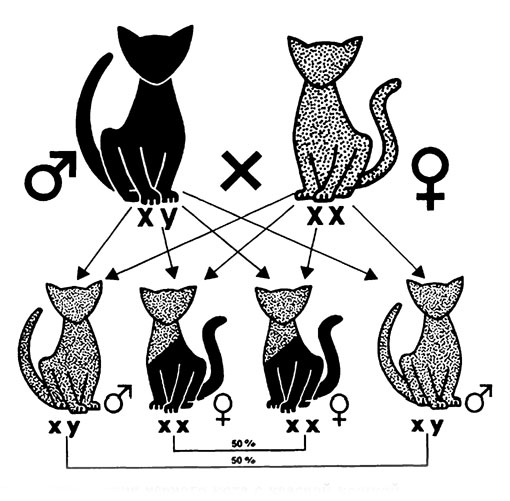
Рис. 17. Схема результатов спаривания черного кота с красной кошкой
Подавляющее большинство генов заложено в ядрах клеток. Носителями генов являются хромосомы. Под микроскопом после соответствующей окраски видно, что хромосомы в метафазе клеточного деления представляют собой линейные образования, количество которых для каждого признака стабильно. Основу хромосомы составляют парные образования, среди которых имеется набор особой пары, так называемые хромосомы пола, или гоносомы, обозначаемые у млекопитающих через X и У. Остальные хромосомы в парах идентичны и называются аутосомами.
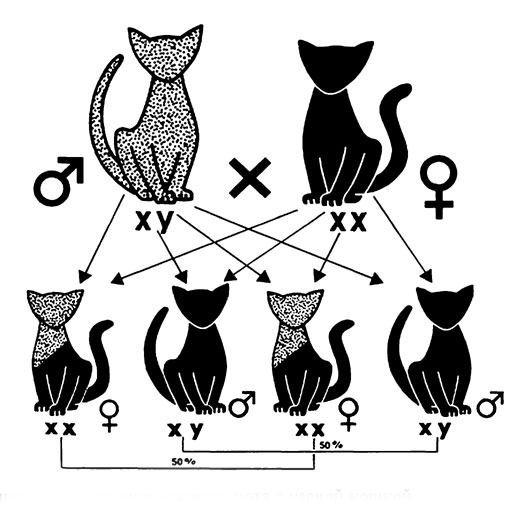
Рис. 18. Схема результатов спаривания красного кота с черной кошкой
Кошка имеет 38 хромосом, причем хромосомы пола кота и кошки отличаются друг от друга: один из полов (кошки) имеет одинаковые половые хромосомы - XX, другой (кот) - разные - X и У.
При созревании половых клеток происходит их редуцированное деление, называемое мейозом, при котором количество хромосом в половых клетках уменьшается вдвое, а при оплодотворении (слиянии яйцеклетки со спермой) в новом зародыше - зиготе - число хромосом удваивается. В зиготе наблюдается обычное количество, причем половина происходит от отца, половина - от матери. Все яйцеклетки содержат хромосому X, а в сперме половина содержит хромосомы X, половина - У, следовательно, пол определяется отцом. То место в хромосоме, где находится ген, получило название локуса.
У диплоидных организмов, к которым относятся и кошки, отдельные гены представлены парой признаков (аллелей), наделенных нарой одинаковых хромосом. Сходство признаков (аллелей) обеих пар называют гомозиготным и обозначают АА, аа. Если эти признаки неодинаковы, такое свойство называют гетерозиготным и обозначают как Аа. Признаки, которые фенотипически проявляются и как гомозиготные, и как гетерозиготные, называют доминантными. Аллели (признаки), которые фенотипически проявляются только в состоянии гомозиготном, называют рецессивными. Совокупность генов ядра или индивида называют генотипом. Внешним выражением генотипа является фенотип, то есть совокупность признаков, характеризующих определенный индивид.
В фенотипе гены подразделяются на:
- гены наиболее активные, так называемые майоргены, или олигогены, которые при образовании признаков практически не подвержены влиянию внешней среди (именно они-то и положены в основу теории Г. Менделя). У кошек, например, такие гены определяют окрас и тин волосяного покрова.
- гены менее активные, так называемые миноргены, или полигены, определяющие возникновение признаков под влиянием внешней среды. У кошек эти гены определяют плодовитость, молочность, рост.
Некоторые признаки обусловлены взаимодействием больше генов неаллельных (локусов), что называется интеракцией (взаимодействием) генов. Превосходство или подчиненность генов при таком взаимодействии носит название эпистаза и гипостаза. У кошек взаимодействие гена А определяет окрас шерсти агути - aguti (однотонный), А генов Ta, T и тb - окрас типа абиссинская, тигровая, мраморная.
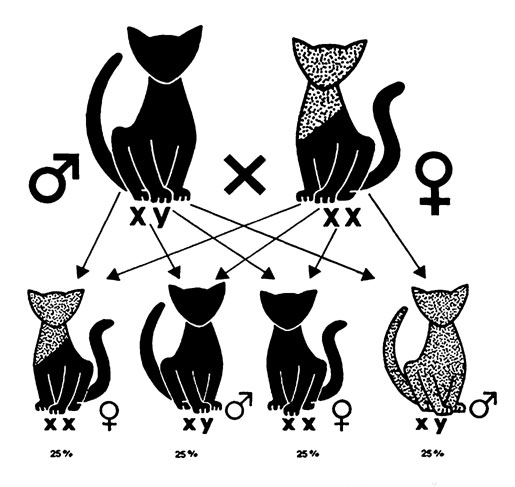
Рис. 19. Схема результатов спаривания черного кота с черепаховой кошкой
Перенос генетической информации происходит в соответствии с определенными закономерностями, главные из которых названы по имени ученых, установивших их, - законы Менделя и Моргана.
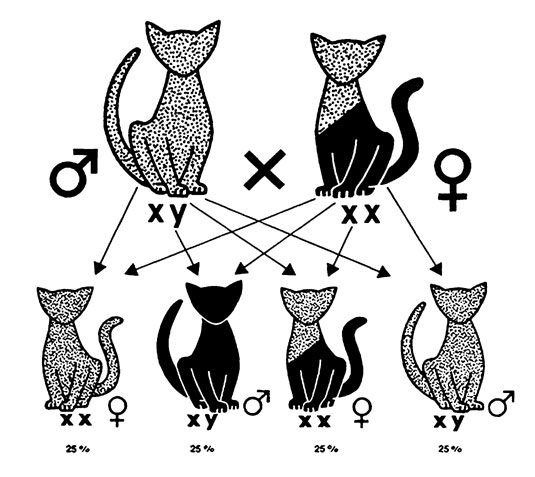
Рис. 20. Схема результатов спаривания красного кота с черепаховой кошкой
Генерация родителей обозначается буквой Р (от латинского слова parentes - родители), а индивиды-производители спариваются по правилам гомозиготных признаков. Потомство, получаемое путем скрещивания индивидов парентальной генерации, обозначается как филиальное (сыновнее) поколение F1(от латинского слова filius - сын). Спариванием индивидов F1 поколений между собой дает генерацию F2 и т. п.
Закон Менделя можно кратко сформулировать следующим образом:
Закономерности однородности F1 поколения, расщепления и идентичности при взаимообратных скрещиваниях. При спаривании раздельных гомозиготных индивидов родительского набора первое (сыновнее поколение) образуется всегда без изменений (униформное). Потомство, полученное при спаривании индивидов F1 поколений, между собой, расщепляется и генотипически, и фенотипически в определенных соотношениях, которые различны в зависимости от того, каково различие признаков у спариваемых индивидов. При спаривании гомозиготов результаты одинаковы, если соответствующий признак имеется у отца или матери. Например, при спаривании гомозиготных белых и черных кошек можно получить белое потомство, если спаривать белую кошку с черным котом илии черную кошку с белым котом.

Результаты окраса шерсти котят при спаривании индивидов с аллелей О или о на хромосоме пола X
Закон чистоты признаков. При спаривании индивидов с доминантными аллелями (признаками) и индивидов с рецессивными признаками эти аллели (признаки) не смешиваются, но рецессивные признаки остаются неизменными, хотя в гетерозиготном состоянии могут проявиться. Но как только они попадают в гомозиготное состояние, они снова проявляются в фенотипе. Например, при спаривании кошки мраморного окраса с гомозиготным котом тигрового окраса все потомство полунеизменными, хотя в гетерозиготном состоянии и не могут проявиться. Значит тигровый окрас, так как признаки тигрового окраса являются доминантным признаком. При взаимном спаривании этих индивидов тигрового окраса F1 поколения между собой получим несколько экземпляров мраморного окраса, как и при спаривании матери мраморного окраса с сыном тигрового окраса. При втором липе спаривания, называемом испытательным (тестаментным), соотношение тигровых и мраморных индивидуумов будет 1:1.
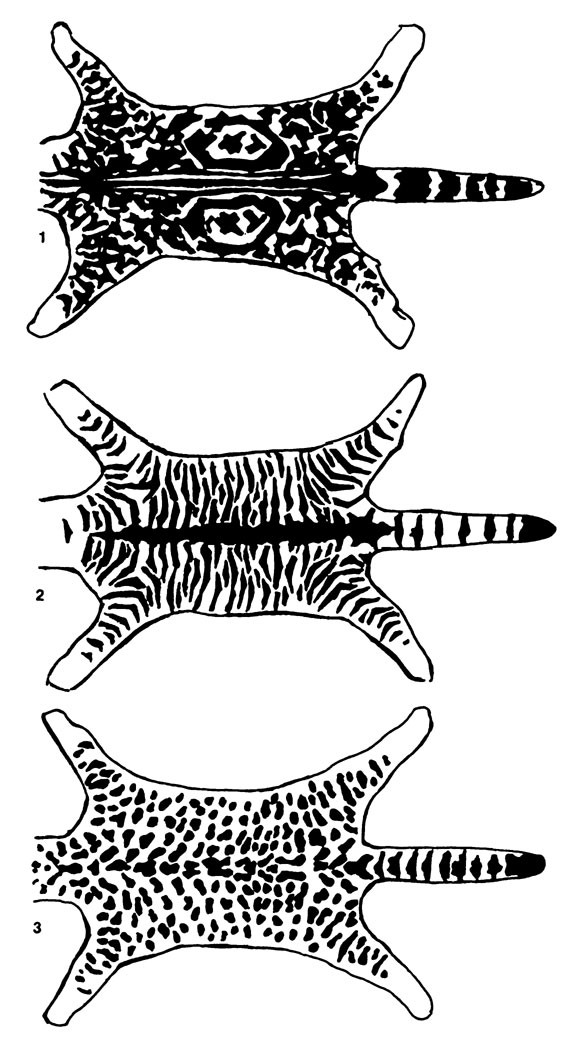
Рис. 21. Образцы рисунка окраса шерсти. 1 - мраморный окрас, 2 - тигровый окрас, 3 - пятнистый (точками) окрас
Закон свободного комбинирования признаков. Если родители отличаются двумя или несколькими парами признаков, эти индивидуальные пары признаков передаются по наследству независимо друг от друга, то есть аллели генов отца и матери комбинируются свободно, без какой-либо закономерности. Морган дополнил данную закономерность: законы вступают в силу в том случае, если каждая пара признаков содержится на своей хромосоме. Такая зависимость дает возможность для большого количества комбинаций у потомства.
У кошек с генетической точки зрения особенно важны следующие признаки: тип, размер (длина), рост, окрас волосяного покрова. Все эти качества обусловлены генами, взаимодействие которых происходит по закону Менделя.
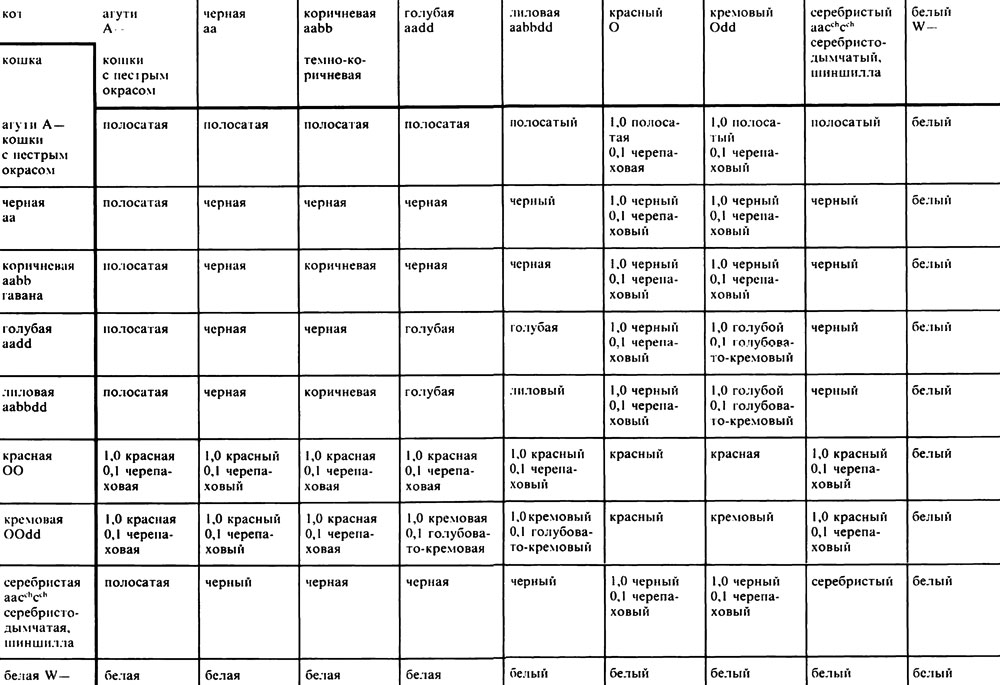
Результаты окраса шерсти при спаривании чистокровных кошек
Чтобы пользоваться единой генетической терминологией, в 1968 г. разработаны Стандарты генетической номенклатуры для домашних кошек (СГНДК), где приведены обозначения для генов, определяющих окрас волосяного покрова, тип кошки и некоторые другие признаки. В 1971 г. в них внес дополнения Робинсон. В такой редакции эти стандарты представлены в настоящей книге. Отдельные признаки характеризуются следующим:
- Ген а - нон агути (non aguti) влияет на единый окрас волосяного покрова без вкраплений или примесей. Этот ген является стойким но отношению к генам Ta, T и tb.
- Ген А - агути (aguti) определяет неравномерность пигментации шерсти, то есть в окрасе волосяного покрова наблюдаются темные и светлые участки. В некоторых случаях темный окрас заметен только на конце волосков. Данный ген взаимодействует с генами Ta, T и tb.
- Ген b - определяет окрас у кошек однотонной окраски, коричневый или шоколадный: у сиамских кошек - коричневый или шоколадный цвет.
- Ген cch - серебристый; определяет у домашних кошек серебристый окрас шерсти, у чистопородных кошек в комбинации с другими генами определяет также дымчатый" шиншилла и камея.
- Ген сh - определяет окрас на определенных местах, т. н. бирманский ("бармский") окрас, типичный для бирманских кошек - голова, конечности и хвост темнее, чем окрас тела.
- Ген сs - сиамский окрас, определяет т. н. акромеланнческий окрас - окрас определенных термальных участков тела, типичный для сиамских кошек.
- Ген с - признак альбиносности, определяет белый окрас. Настоящие альбиносы отличаются не только белым окрасом шерсти, но и красным цветом глаз.
- Ген d - признак разреженного голубого окраса, определяет фенотипичное проявление темного серо-голубого цвета, характерного, например, для британской голубой кошки, для картузской и русской голубой кошек. Этот ген определяет также разреженно-черные цвета у голубовато-полосатых и голубовато-черепаховых кошек. У кошек красного окраса этот ген определяет появление кремового оттенка в окрасе. У сиамских кошек ген d определяет голубой окрас.
- Ген h - признак безволосого покрова, определяет бедность волосяного покрова на коже, которая бывает почти лысой, толстой и покрыта шерстью (очесами) только в местах кожных сгибов.
- Ген l - типичен для персидских длинношерстных кошек, определяет образование длинного и тонкого волосяного покрова с густым подшерстком.
- Ген М - типичен для мэнских бесхвостых кошек (Великобритания), определяет отсутствие хвоста у мэнских кошек, отличающихся характерным строением задней части тела.
- Ген О - признак оранжевого (красного) окраса, локализованного в хромосоме иола X: этот тип окраса зависит от иола особи. Интенсивность оранжево-рыжего окраса весьма разнообразна и определяется также другими генами. Если ген О имеется в обеих хромосомах у кошки-самки, то появляется окрас красный (оранжевый); когда же ген О присутствует только в одной хромосоме X, образуется черепаховый окрас. Если его нет ни в одной из хромосом X, образуется черный окрас. У котов присутствие гена О в хромосоме X вызывает красноватый окрас, при его отсутствии - черный. Так как у котов обычно одна хромосома X, а другая - Y, у них не бывает черепахового окраса. Такой редкий окрас может иногда появиться и у кота, но такие индивиды считаются по экстерьеру промежуточными (интерсексы), у них лишь одна хромосома X (2n = 39 XXY); они бесплодны.
- Гены r, re, rg, ro определяют волнистость шерсти у различных пород кошек рекс. Эти гены индивидуально специфичны: при взаимном скрещивании отдельных племен этих кошек рекс не появляется промежуточный тип кошки "рексового" волнистого, с очесами волосяного покрова: отмечаются индивиды с нормальной шерстью.
- Ген S - белая пятнистость - определяет образование белых пятен в окрасе волосяного покрова, причем соотношение белого и другого цвета на поверхности тела может варьировать: имеются индивиды с едва заметными пятнышками, с несколькими белыми пятнами средних размеров и белыми пятнами, занимающими значительную часть поверхности волосяного покрова. Величина этих пятен определяется другими генами. У чистопородных кошек этот ген дает белую окраску у кошек с двухцветным окрасом (биколор) и у кошек с белесо-черепаховым окрасом (трехцветная окраска).
- Ген Та - абиссинский окрас, определяет абиссинский окрас кошек, при котором на отдельных волосках встречаются светлые и темные участки, а кончик волоска темный, что типично для меланжевой окраски. Среди чистопородных особей такой окрас встречается у абиссинских и сомалийских кошек.
- Ген Т - тигровый окрас, определяет характерный окрас шерсти - полосатый: в виде полос, расположенных вертикально от спины до брюшной полости. Эти полосы темнее основного тона, иногда черные или темно-красные (у красных тигровых видов). Данный ген является гипостатичным по отношению к генам Тa, но доминантным по отношению к гену tb. Модифицированной формой тигрового окраса является крапинка, определяемая другими генами независимо от генов Т и tb.
- Ген tb - мраморный окрас, определяет мраморный рисунок на основном тоне окраса: выступает обычно в форме неправильных меандров (см. рис.), которые расположены несимметрично по обеим сторонам туловища. Окрас такого типа появляется только у гомозиготов, причем разной формы, размещается на различных участках волосяного покрова, но основной окрас не должен быть белым.
- Ген w - доминантный белый окрас, определяет белый окрас шерсти у кошек неальбиносов. При комбинации доминантного белого окраса с голубыми глазами у некоторых кошек наблюдаются нарушения состояния здоровья (глухота, стерильность котов и под.), что обусловлено, очевидно, генами, взаимосвязанными с генами, в которых белый окрас и голубоглазость являются доминирующим признаком.
- Ген Wh - признак волнистости шерсти, определяет волнистую шерсть у кошек.
| Аллели (признаки) генов по СГНДК | Соответствующие противоположные признаки |
| а - нон агути = одинаковый окрас по всей длине волоса | А - агути = окрас |
| b - коричневый окрас | В - неразреженный цвет окраса |
| С - однотонный окрас | * |
| cch - серебристый разреженный окрас | S - однотонный окрас |
| сh - бирманский ("бармский") окрас | С - однотонный окрас |
| сs - "сиамский" окрас | С - однотонный окрас |
| с - альбиносы | С - однотонный окрас |
| d - голубой разреженный окрас | D - неразреженный окрас |
| h - безволосый кожный покров | Н - нормальный кожный покров |
| l - персидский длинношерстный волосяной покров | L - нормальная длина волосяного покрова |
| М - отсутствие хвоста (мэнские кошки) | * |
| О - оранжевый окрас (зависит от пола) | о - черный окрас |
| r - корнрекс | R - нормальная шерсть |
| re - шерсть девонрекс | R - нормальная шерсть |
| rg - немецкий рекс | R - нормальная шерсть |
| ro - шерсть т. н. "орегон-рекс" | R - нормальная шерсть |
| S - белая пятнистость на волосяном покрове | S - отсутствие белых пятен на волосяном покрове |
| Тa - абиссинский окрас | * |
| Т - тигровый окрас | * |
| tb - мраморный окрас | * |
| W - доминантный белый окрас | w - окрас не белого, иного цвета |
| Wh - волнистость шерсти | wh - нормальная шерсть |
Группировка отдельных аллелей (признаков), определяющих тип и окрас шерсти, создает генотипы пород кошек, которые проявляются в их фенотипе - иными словами, в действительности существующем типе и окрасе шерсти. В качестве примера можно привести несколько случайно выбранных пород чистопородных кошек, генотип которых отражает наследственность типа и окраса шерсти; аналогичным образом это можно определить и у любых других пород. При описании генотипов необходимо помнить, что некоторые стабильные доминантные признаки (напр., LL - нормальная длина шерсти, АА - агутный окрас и т. д.) в генотипах не упоминаются, когда очевидно, что они отсутствуют. Приведем примеры:
| * | генотип |
| европейская короткошерстная кошка, окрас белесо-черепаховый | aaOoS- |
| белая европейская короткошерстная кошка | W- |
| картузская кошка | aadd |
| сиамская кошка с лиловыми отметинами | cscsbbdd |
| персидская красно-белая кошка (биколор) | llO-S- |
| персидская дымчатая кошка | llaacchcch |
| персидская голубовато-кремовая кошка | llOodd |
| коричневая мраморная европейская короткошерстная кошка | tbtb |
При содержании чистопородных кошек знание генетики необходимо для выведения редких по экстерьеру индивидов или для выведения новых пород. Надо постоянно помнить о законах генетики и для спаривания отбирать особи среди отдельных пород с учетом соответствующего окраса, чтобы получить индивид, отвечающий стандарту. Знание основных законов генетики позволяет специалистам точно определить, является ли тот или иной индивид (в зависимости от окраса шерсти) действительно потомком определенной родительской пары и избежать различного рода подлогов.
В данной книге невозможно подробно описать все типы спаривания и генетических комбинаций, но для контроля специалистам предлагаются следующие таблицы окраса индивидов при спаривании чистопородных кошек. Там, где имеется больше возможностей (например, при спаривании кошек черепахового окраса с красными и черными котами), при малом количестве котят отмечают и некоторые случайные явления: наличие одноцветности в окрасе у всех котят в помете, если они одного пола.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://mur-r.ru/ 'Библиотека о кошачьих'